Сельское хозяйство
Экологические условия обитания бентосных гидробионтов на участке «Северный»
Лента новостей
Помощь за 5 секунд: пользователям каршеринга стала доступна приоритетная линия в случае ДТП
Фонд «Полилог» запускает федеральный просветительский проект «Вестник мецената»
Банк Уралсиб запустил программу автокредитования на покупку автомобиля с пробегом у физлиц на Авито
SMS Traffic готова противостоять новым вызовам в сфере киберугроз
Акватория расположена в западном районе Северного Каспия. Южная его граница проходит южнее Мангышлакского порога, захватывая часть Среднего Каспия. За условную границу между Северным и Средним Каспием принимают линию, соединяющую о. Чечень и м. Тюб-Караган.[1]
Площадь западной части, при современном уровне моря — 27,0 м абс., составляет 26 тыс. км2 или около 34% всего Северного Каспия, а лицензионного участка «Тюлений» -7,6 тыс. км2
или около 17% площади западной части. Если площадь с глубинами более 5 м в Северном Каспии занимает лишь 32%, то на лицензионном участке, который расположен в приглубой зоне, она составляет около 94% его акватории. В целом, на лицензионном участке глубины изменяются от 3 м на севере акватории до 36 м на юго-востоке.
При общей выравненности рельефа дна здесь наблюдаются и некоторые неровности в виде положительных и отрицательных форм рельефа. К первым относятся банки и острова, ко вторым – бороздины.
На лицензионном участке расположены Кулалинская и Безымянная банки, являющиеся частью Мангышлакского порога. На прилегающей к участку акватории находятся Большая Жемчужная, Чеченская, Становая, Песчаная и др.В непосредственной близости расположена группа островов аккумулятивного происхождения: Чечень, Тюлений, Чистая банка, Укатный, Кулалы и др.
Обращает на себя внимание морфологическое сходство банок и островов. Обычно они состоят из одного, а чаще из нескольких дугообразно изогнутых валов – подводных или надводных баров. Наличие баров и рыхлый состав наносов, преимущественно песок и ракуша, слагающие банки и острова Северного Каспия, подтверждают мнение об их волновом генезисе. Однако их зональное расположение обусловлено особенностями геологической структуры (Леонтьев, 1969).
Преобладающими типами донных отложений являются мелкая ракуша и крупный алеврит, на прилегающей к нему акватории – крупный алеврит, мелкий песок и мелкая ракуша.
Гидролого-гидрохимический режим Северного Каспия отличается значительной сезонной и межгодовой изменчивостью, определяемой в основном стоком Волги и водообменом со Средним Каспием.
Для северной части моря характерны существенные изменения природных условий, обусловленные комплексом климатических, гидрологических и геологических процессов, характерных для бассейна. Среди компонентов природного комплекса наиболее динамичны морфометрия и топография водоема.
Так, с 1978 г. по 1995 г., в период трансгрессии, уровень Каспия повысился с -28,92 до -26,54 м абс. (по в/п Баку), т.е. на 238 см. В результате этого площадь Северного Каспия возросла до 109,2 тыс. км2, а объем его водных масс- до 541 км3 или соответственно в 1,5 и 1,7 раза. Несколько изменилась конфигурация береговой линии. Оказались подтопленными крупные острова и затоплены мелкие. К 2007 г. уровень моря понизился примерно на 0,5 м и его среднее годовое значение в 2006 г. составило -27,0 м абс. [1, 2, 3]
Северный Каспий в той или иной степени замерзает ежегодно. Значительная его акватория покрывается неподвижным льдом — припаем, который составляет основную часть ледового покрова. Площадь под плавучим льдом намного меньше. Плавучий лед имеет наибольшее распространение в марте-апреле при общем взломе припая. Полоса плавучего льда шириной 10-20 миль обычно окаймляет границы припая.
Ледовый покров препятствует нормальному судоходству и может нанести значительный ущерб нефтегазодобывающей отрасли.
Обычно первое появление льда происходит в середине ноября в северо-восточной части моря. К концу месяца льдообразование быстро распространяется по акватории, охватывая северное побережье, включая приустьевое взморье Волги. В первой декаде декабря лед появляется во всех мелководных районах Северного Каспия, ограниченных трех — четырехметровыми изобатами. К концу месяца льдообразование распространяется в более мористые районы. Припай получает наибольшее распространение в январе, когда его граница проходит от о. Чечень к о. Тюлений и далее по 5-метровой изобате к о. Кулалы и п-ову Тюб-Караган. В суровые зимы граница припая проходит по 10-20 метровым глубинам, а максимальная ледовитость моря отмечается в феврале. В такие зимы толщина льда на северо-востоке моря достигает 80-90 см, в центральных и юго-западных районах- 50-60 см (Валлер, 1970).
В северной части моря вследствие мелководности образуются многочисленные стамухи и барьеры стамух. Последние могут достигать более полутора километров в длину, десятков метров в ширину и 10–12 м в высоту.
В умеренные зимы в первой декаде марта в центральных районах Северного Каспия, во второй декаде – на отмелой устьевой части Волги припай окончательно разрушается и превращается в плавучий лед, состоящий из ледяных полей и битого льда, которые дрейфуют и уменьшаются в размерах. Одновременно происходит и термическое разрушение льда. Полное очищение Северного Каспия ото льда южнее линии о. Чечень — о. Кулалы происходит в конце первой декады марта, южнее линии о. Тюлений — в конце второй декады, по линии о. Чистая банка — о. Укатный – в конце марта. Окончательное очищение приустьевого взморья Волги наблюдается в первой декаде апреля (Сокольский и д.р. 2014).
На основе анализа многолетних материалов можно сделать вывод, что ледовый режим на акватории участка «Тюлений» наиболее динамичен на всей акватории Северного Каспия, характеризуется активным движением ледовых полей под воздействием нагонных явлений из средней части моря. Для участка характерно интенсивное торошение льда и максимальное для Северного Каспия воздействие ледовых нагрузок.
Температура воды
Архипова (1955) в Северном Каспии выделила 7 физико-географических районов, отличающихся формированием термического режима под воздействием климатических факторов, географического положения, батиметрических особенностей и рельефа дна, притока речных вод. Каждый из этих районов характеризуется однородностью внешнего и внутреннего теплооборотов.
Северная часть исследуемого участка входит в состав приустьевого (волжского) района, южная – центрального (глубоководного). Для первого района характерно наименьшее поступление солнечного тепла (115 ккал/см2
в год), для второго (южного) – несколько большая (119 ккал/см2 в год).
Из элементов внутреннего теплообмена для северной зоны участка следует выделить адвекцию вод из отмелого устьевого взморья р. Волги, для южной – из Среднего Каспия. В северной зоне доля внутреннего теплооборота невелика по сравнению с внешними формируемыми (климатическими) факторами, в южной — определяющим служит внутренний теплооборот.
По совокупности факторов северная часть участка используется ихтиофауной в качестве нерестового, нагульного и миграционного ареалов; южная – преимущественно в качестве нагульных преднерестовых и предзимовальных миграционных концентраций рыб.
В целом, оценивая температурный режим участка «Тюлений», следует признать его неоднородным, с резким контрастом внутригодовых изменений между его северной и южной частями.
Для относительно мелководной северной части акватории с глубинами до 5-8 м с началом весеннего прогрева (апрель) характерно формирование вертикальной структуры с небольшими температурными градиентами, которые в южной части участка в это время сглажены.
В начале летнего периода (июнь) вследствие интенсивного прогрева на акватории, ограниченной глубинами 10 м, развивается гомотермия; в южной части участка вертикальные различия складываются в условиях прогрева поверхностных вод и усиления компенсационного подтока из Среднего Каспия. Термоклин начинает формироваться во второй половине мая, ограничивая вертикальное перемешивание водных масс. Температурный максимум в это время формируется в северной, сравнительно мелководной части акватории.
Осенью вертикальные температурные различия нивелируются в результате сезонного выхолаживания поверхностных вод, разрушения термоклина и перемешивания водных масс. При этом сохраняется повышенный теплозапас в южной приглубой части участка.
За период весенних исследований наблюдались заметные различия в прогреве вод поверхностного и придонного горизонтов (табл. 1.1).
Максимальная разница температурных величин поверхностного и придонного горизонтов характерна для южных районов участка, где на фоне сравнительно высокого прогрева поверхностных вод (13-15оС) температура в придонном слое в среднем составляла 7-10оС вследствие адвекции холодных среднекаспийских водных масс.
Таблица 1.1
Средние величины температуры воды, оС
Пространственное распределение температуры в поверхностном горизонте по осредненным за рассматриваемый период данным характеризуется более высокими величинами в северной и центральной частях участка; более низкое теплосодержание в северо-западной и западной частях обусловлено выносом сравнительно холодных волжских вод из Волго-Каспийского канала. В придонном горизонте четко прослеживается поступление вод из Среднего Каспия, в результате чего южные и юго-восточные районы рассматриваемой акватории заняты холодными (от 9 до 7,2 оС) водами.
Летом при интенсивном прогреве поверхностных вод и высоком теплообмене с придонными водами вертикальные различия сглаживаются. Наиболее заметно это происходит на участках в пределах 10-метровой изобаты. Южнее разница температур воды поверхностного и придонного горизонтов на фоне усиления в начале лета компенсационного подтока из Среднего Каспия составляла 8-9оС. Температурный минимум у дна достигал в среднем 9,4оС
Температурный режим в северной и центральной частях участка в это время характеризуется формированием летней гомотермии и минимальными вертикальными градиентами температуры воды между поверхностным (21-23оС) и придонным (19-23оС) горизонтами. В центральной части рассматриваемого участка максимальная температура воды в поверхностном слое в летнее время достигала 26,8 оС, в придонном — 26,7оС.
В начале осени сохраняется сравнительно высокий теплозапас. На западе Северного Каспия средняя температура воды поверхностного горизонта в сентябре превысила среднемноголетнюю на 1,2оС, придонного — на 0,8оС. Вследствие этого средние величины температуры воды несколько превышали летние. Вместе с этим для начала осени характерно размывание слоя температурного скачка на фоне начинающегося выхолаживания вод поверхностного горизонта. Температурный максимум (до 25,1оС) в сентябре перемещается в южную часть участка.
Более мелководные северные участки в это время отличаются повышенным теплообменом с атмосферой. В придонном горизонте характерно сохранение особенностей пространственного распределения температуры воды: более высокий прогрев (до 26,6 оС) в северной и центральной частях участка и пониженный – в зоне адвекции среднекаспийских вод (до 8,4 оС).
В конце осени (ноябрь), вертикальные различия температуры воды на основной части лицензионного участка минимальны вследствие перемешивания водных масс (рис. 6.4). Максимальное выхолаживание происходит в его северных мелководных участках (до 2-3 оС); относительно высокий теплозапас сохраняется в юго-восточной части акватории (до 11,8 оС). [1, 5, 6]
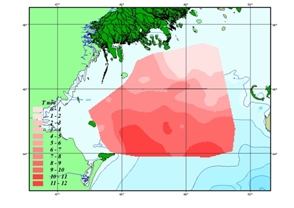
а)
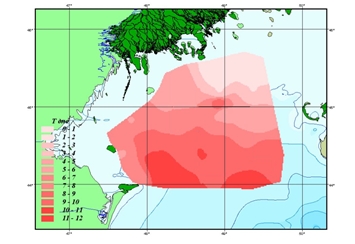
б)
Рис. 1.1 Температура воды, осенний период (ноябрь)
а) поверхностный горизонт; б) придонный горизонт, оС
В целом, для рассматриваемого участка характерны основные особенности температурного режима западной части Северного Каспия. Сезонная динамика отличается интенсивным прогревом в июне, который в середине летнего периода достигает максимума. Вместе с этим, в начале осени характерно сохранение сравнительно высоких температурных величин. Повышенный теплозапас в большей степени характерен для южных участков акватории. Вертикальное распределение температуры воды определяется прогревом вод поверхностного горизонта и поступлением среднекаспийских более холодных вод. При этом начало образования температурной стратификации характерно для окончания весеннего периода при сравнительно высоком прогреве поверхностных вод. Летом, в условиях высокого прогрева, на основной части акватории (преимущественно до 10-метровой изобаты), занимаемой лицензионным участком, формируются однородные по вертикали водные массы; температурная стратификация сохраняется в его южной части.
В северо-западной и западной частях полигона прослеживается связь между типами донных отложений и прозрачностью воды. В совокупности они влияют на развитие и распределение макрофито- и зообентоса. По мере продвижения на юг и восток доля илистых отложений уменьшается, а прозрачность воды увеличивается. Так, на илисто-песчаных грунтах она составляет 1,5-2,0 м; на песчаных — 2,0-3,0 м. В центре полигона и в южной приглубой его части, где поверхность дна в основном представлена ракушей, прозрачность доходит до 3,0-6,0 м. Благодаря большому количеству организмов-фильтраторов и поступлению среднекаспийских вод на юго-востоке прозрачность воды достигает 8 м.
В июне, в период прохождения волны половодья в море, в южной части полигона в толще воды отмечено наличие большого количества крупной взвеси (Ушивцев, Чиженкова, 2004). Длина отдельных нитей может достигать 10 см и более. Под слоем термоклина взвесь не встречается, а прозрачность воды значительно возрастает — в 1,5-2 раза.
Уровень токсичности воды на участке «Тюлений» по результатам биотестирования
В условиях растущего техногенного загрязнения окружающей среды, в частности природных водных объектов, оценка их состояния в значительной мере зависит от достоверности и оперативности контроля качества воды. Вместе с тем даже полный физико-химический анализ воды по всем установленным нормативам не позволяет оценить результаты комплексного воздействия присутствующих в воде загрязняющих веществ на экосистему водного объекта в целом. В связи с остротой современной ситуации в отношении химического загрязнения водоемов необходимы не только всесторонние исследования биологического действия и последствий антропогенного нарушения состава морской среды, но и широкое использование токсикологических методов в практике предотвращения загрязнения. К числу наиболее эффективных мероприятий, отвечающих целям охранных мер, учитывающих экологический аспект, относится применение для оценки токсичности воды методов биотестирования с использованием чувствительных тест-организмов. Биотестирование позволяет получить интегральную оценку токсичности воды и значительно сократить объем химико-аналитической работы, повысить оперативность контроля над состоянием природных водных объектов.
Результаты биотестирования исследуемых сред позволяют оперативно оценивать воздействие на жизнедеятельность водных организмов различных загрязнений, причем не отдельных токсикантов, а их соединений, частью неизвестной природы и не выявляемых другими методами анализа. Определение полного перечня присутствующих в исследуемых водах веществ современными аналитическими методами связано с большими материальными затратами. Выявление достоверного количественного значения тест-параметров (смертность, выживаемость, плодовитость) и тест-реакций организма (изменение морфологического, биохимического, поведенческого и функционального показателей) позволяет получить наиболее полные сведения при минимальных затратах на выполнение контрольных операций.
Исследования в области разработки и использования метода биотестирования в водоохранной практике проводились во многих научно-исследовательских и учебных институтах. В 1980 г. была признана необходимость применения биотестирования как показателя оперативной интегральной диагностики качества вод. В 1981-1986 гг. методики биотестирования были апробированы и рекомендованы для определения токсичности сточных и природных вод.
Определение уровня токсичности морских вод участка «Северный» методом биотестирования проводилось согласно Руководству, утвержденному Министерством природных ресурсов РФ (Руководство по определению …, 2002; Методические указания …, 1978). Руководство подготовлено Центром Российского регистра гидротехнических сооружений и государственного водного кадастра МПР России совместно со специалистами ВНИРО Госкомрыболовства России и УНИИ экологических проблем Минэкоресурсов Украины. В основу предлагаемой системы морских токсикологических биотестов положены результаты обобщения многолетних экспериментальных исследований ВНИРО по проблеме загрязнения водоемов и многочисленных литературных данных, позволивших выявить особенности реагирования гидробионтов разных систематических групп на токсические примеси различной природы и происхождения. Экспериментальная часть исследований проводилась на культуре морских одноклеточных водорослей Phaeodactylum tricornutum, на планктонных ракообразных Acartia tonsa, личинках хирономид Chironomus gr. salinarius и молоди гуппи Poecillia reticulata Peters.